Reforzamiento del aislamiento reproductivo en el contacto secundario, especiación y desplazamiento de caracteres
Hola a todos

Siguiendo al post anterior, el refuerzo o reforzamiento ocurre en el contacto secundario entre poblaciones próximas, esto es representado cuando las poblaciones biológicas presentan comúnmente un aislamiento potscigótico, con híbridos de baja eficacia biológica, apareciendo una barrera precigótica favoreciendo la especiación. El modelo de selección natural es el responsable de la aparición del aislamiento precigótico, actúa como refuerzo en regiones de contacto entre las poblaciones, este ejerce una presión en contra de los híbridos, con una baja viabilidad de reproducción.
Una de las discusiones para abordar la propuesta del reforzamiento entre las poblaciones biológicas, es comprender el grado o nivel de barrera reproductiva entre las especies simpátricas y alopátricas, siendo las más estudiadas experimentalmente. A pesar de todo, las evidencias para respaldar el refuerzo precigótico, estas mismas pueden ser explicadas por otros mecanismos o procesos evolutivos. Por ejemplo, el desplazamiento de caracteres.
Especiación simpátrica y Desplazamiento de caracteres
Antes de continuar es imprescindible esclarecer dos conceptos. El primero de ellos es la especiación simpátrica, que en otro post voy a ampliar la explicación, sin embargo como es un mecanismo que estoy introduciendo es conveniente aclarar su concepto. Este tipo de especiación, ocurre cuando en una misma región geográfica los individuos de una población biológica comienzan a ocupar diferentes nichos ecológicos a tal punto que forman barreras reproductivas, algunos consideran aspectos ecológicos así como etológicos para el desarrollo de la divergencia evolutiva.
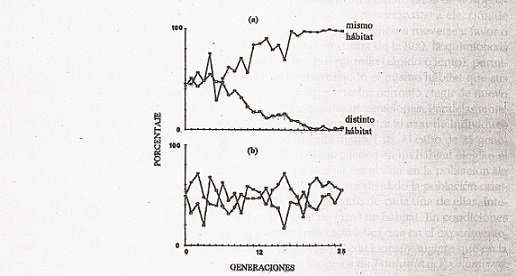
Estudio de casos: Un experimento con la población biológica de la especie Drosophila melanogaster conocida como la mosca de la fruta sigue así: En un hábitat elaborado en un laboratorio, cada especie podía elegir el microhábitat que deseaba, estos espacios estaban controladas determinadas variables como la incidencia de luz, el tiempo para desarrollarse y otros. Además, las primeras generaciones de hembras fueron controladas, debido que solo podían reproducirse en el mismo hábitat seleccionado por sus progenitores. Después de varias descendencias, la prole una vez más podía elegir el hábitat que consideraba de su preferencia, siguiendo este patrón durante numerosas generaciones. A la par, fueron tomadas una pequeña parte de los descendientes. Al cabo de veinticinco generaciones filiales, obtuvieron como resultado que al menos el dos por ciento, seleccionaba un microhábitat diferente de donde residen sus progenitores, y el resto escogía al azar uno de los habitas controlados. Así pues, como podemos ver en el gráfico en una misma población biológica, existen individuos que tienden a preferir un hábitat distinto a otros, pudiendo llegar al punto que impida el flujo genético, y dividiendo a la población en dos o más grupos a tal punto que se consideren especies diferentes (según el concepto de especie biológico).

El segundo es el desplazamiento de caracteres. Cuando se presentan escenarios en la naturaleza, en el cual hay competencia de dos o más poblaciones biológicas ubicadas comúnmente en un mismo espacio geográfico, tienden a presentar rasgos fenotípicos diferentes para reducir la competencia por algún recurso. Esta competencia, es una interacción que puede ser con la misma especie o con otras especies. De igual manera, dentro de la misma población biológica, podemos tener individuos con rasgos pequeños o mutaciones que podrían ser importantes durante la interacción por obtener el recurso a tal punto que marcaría un cambio evolutivo.
El desplazamiento de rasgos o caracteres asume que la competencia por algo, bien sea un alimento o apareamiento, tenderá a beneficiar a una selección de rasgos distintos. Por ello, es común que una misma región geográfica es posible observar más variedades morfológicas que en poblaciones separadas geográficamente, es decir, en poblaciones simpátricas es mayor la diferenciación que en poblaciones alopátricas.
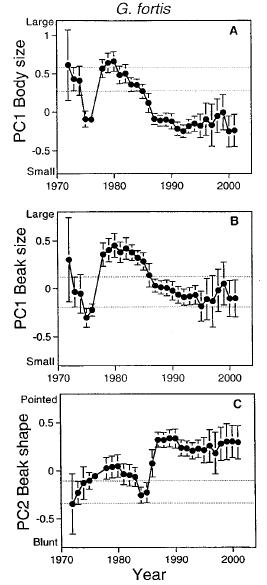
Estudio de casos: Tenemos los estudios realizados en los Galápagos (o archipiélagos galápagos). En esta región geográfica encontramos espacios geográficos distribuidos y separadas por agua. Cuando fueron estudiados los pinzones de Darwin, notaron que según las interacciones con el hábitat (condiciones dentro del ambiente, las épocas o estaciones del año, tipo de alimentación, relación con otras aves y demás) y su implicaciones genéticas, ocurría un desplazamiento de carácter dentro de una misma especie. Ejemplo de ello, la especie Geospiza fortis durante un tiempo de sequía estuvo compitiendo con la especie Geospiza fuliginosa, ambas están en competencia por el recurso alimenticio.

Durante un año de sequía la población G. fortis se vio afectada, puesto que no todos los individuos estaban adaptados a comer semillas que estuviesen un pequeño tamaño, sin embargo, ambas especies tienen individuos con rasgos (en menor proporción) que no se interponen con la alimentación de la otra. Por lo cual, estos rasgos fenotípicos que residen en sus genes fueron mostrándose en mayor proporción durante varias generaciones. Como vemos en la figura estamos en presencia de individuos, de una misma población biológica, que presentan un desplazamiento caracteres durante el transcurso de los años, por el recurso alimenticio. Vale destacar, que para que se de esto, deben ocurrir algunas condiciones y una de ellas es que posean variaciones alélicas con estos caracteres. Además, esto señala que estas poblaciones pueden coexistir en mismo hábitat, a pesar de competir por un recurso que a veces puede ser limitado.
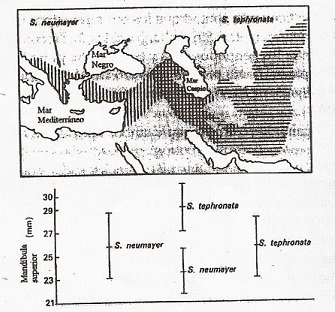
A propósito de lo mencionado, que en espacios de poblaciones simpátricas es posible observar mayor diferenciación fenotípica en poblaciones alopátricas. En aves que están presenten en la misma región, expresan una diferencia morfológica de picos que especies que se encuentran separadas geográficamente. Como podemos ver en la imagen, las poblaciones de Sitta neumayer y Sitta tephronata, están aisladas geográficamente y presentan longitudes de pico similares, mientras que un grupo de esta comunidad biológica, cuando están en un mismo territorio presentan longitudes de pico diferentes. Vale decir, que esto es empleado para el caso del recurso alimenticio, los mismo es plausible aplicar si hablamos de la selección sexual.

Modelos teóricas para corroborar la veracidad del reforzamiento
Es imprescindible contar con dos procesos para sustentar teóricamente el reforzamiento, el primero es el flujo de genes; La repercusión del intercambio de los genes tiene su importancia en las zonas de contacto donde puede existir un cambio gradual en la especie (clina), cuando es estable la hibridación, donde la barrera reproductiva no se ha completado. En otras palabras, presentamos una zona denominada zona de contacto, en la cual dos o más individuos se reproducen y forman híbridos, estos dos grupos no han completado sus aislamiento reproductivo. Uno, debido a la presión selectiva en contra de la formación de híbridos, y dos, a individuos que están próximos a la zona de contacto que no reproducen híbridos.
El segundo proceso es la recombinación, un modelo sencillo y esquematizado sigue así;
- El locus Z implica una barrera poscigótica.
- El heterocigoto Z1Z2 presenta un menor nivel de aptitud.
- El locus X implica la preferencia para aparearse.
Ahora bien, supongamos que tenemos dos grupos que tienen un nivel de aislamiento postzigótico, el primero con sus genes Z1Z1X1X1 y el segundo Z2Z2X1X1. Entonces, aparece una mutación X2, la cual está implicada en la preferencia para aparearse, y que además está ligada al grupo con el genotipo Z2Z2, en consiguiente la frecuencia de X2 podría ir en aumento en este grupo, es decir el X2 se va distribuir en las especies con el genotipo Z2Z2.
- Estamos en presencia de dos tipos de preferencia para seleccionar a individuos para aparearse, X1 y X2.
- Mientras que los genotipos X1 se expanden en los grupos con Z1, los X2 se expanden con los de Z2.
De tal manera, a medida que se expande estas preferencias, se formaran dos grupos dentro de la población para escoger de diferente pareja para reproducirse. Así pues, los grupos que están en la zona de contacto con un leve aislamiento postcigoto se refuerza con un tipo de aislamiento precigotico. Sin embargo, puede ocurrir un escenario donde los locus se recombinen, dando como resultado que la expansión del nuevo gen X2 se distribuya en toda la población, por consiguiente la selección no es favorecida. No obstante, este modelo se vuelve más complejo porque sabemos que en los estudios genéticos no son solo dos.

Fuente de las imágenes: A B C D E
Referencia de la información:
- Fontdevila y Moya. EVOLUCIÓN: ORIGEN, ADAPTACIÓN Y DIVERGENCIA DE LAS ESPECIES
- Perfectti. ESPECIACIÓN: MODOS Y MECANISMOS
- Resource Partitioning and Why It Matters
- Pinzones de Darwin y Selección Natural. Evolución a tiempo real
Excelente trabajo amigo como siempre destacándose y enseñándonos estas lecciones que son muy instructivo @hogarcosmico. Saludos ; )
Gracias por apreciar el trabajo. Feliz día. :D